Phaeocystis
Phaeocystis ist eine Gattung von Algen, die zur Klasse der Prymnesiophyten (Prymnesiophyceae) und damit zum großen Phylum (Abteilung) der Haptophyten (Haptophyta) gehört.[1] Phaeocystis-Arten sind im marinen Phytoplankton weit verbreitet und können in einem weiten Temperaturbereich („eurythermal“) und Salzgehaltsbereich („euryhalin“) vorkommen.[2] Mitglieder dieser Gattung leben sowohl im offenen Ozean als auch im Meereis.[3]
| Phaeocystis | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|

Phaeocystis antarctica: Zelle in Zellkultur. | ||||||||||
| Systematik | ||||||||||
| ||||||||||
| Wissenschaftlicher Name | ||||||||||
| Phaeocystis | ||||||||||
| Lagerheim, 1893 |


.jpg)
Phaeocystis-Arten haben einen vielgestaltigen (polymorphen) Lebenszyklus, der von freilebenden Zellen bis zu großen Kolonien reicht.[2] Auf diese Weise sind einige Arten von Phaecocystis in der Lage, schädliche Algenblüten (en. harmful algal blooms, HABs) zu bilden.
Arten
Bearbeiten

Gattung Phaeocystis[5][6][7][8]
- P. amoeboidea Büttner, 1910 (Kieler Bucht, Holtenauer Ufer)
- P. antarctica Karsten, 1905 (Atlantischer Sektor der antarktischen zirkumpolaren Konvergenz)
- P. brucei Mangin, 1922 (saisonale Eiszone – seasonal ice zone, SIZ – des Südlichen Ozeans)
- P. cordata A. Zingone & M. J. Chrétiennot-Dinet, 2000 (Mittelmeer)
- P. globosa Scherffel, 1899 (Helgoland)
- P. jahnii A. Zingone, 2000 (Mittelmeer)
- P. pouchetii (Hariot) Lagerheim, 1896 (Typus / Holotyp Spezies, Nordatlantik: entlang der Küste Norwegens von den Lofoten bis zum Varangerfjord)
- P. rex Andersen, Bailey, Decelle & Probert, 2015 (Arabisches Meer, nicht gelistet bei WoRMS)[9]
- P. scrobiculata Ø. Moestrup, 1979 (nicht in Kultur, Neuseeland)
- P. sphaeroides Büttner, 1910 (Kieler Bucht, Möltenorter Ufer)
- „P. sp. AC618“ (nicht-klassifizierter Kandidat nach NCBI)
- „P. sp. Hongkong2“ (nicht-klassifizierter Kandidat nach NCBI)
- „P. sp. PLY559“ (nicht-klassifizierter Kandidat nach NCBI)
- „P. sp. Hongkong2“ (nicht-klassifizierter Kandidat nach NCBI)
- „P. sp. Santou“ (nicht-klassifizierter Kandidat nach NCBI)
- „P. sp. SIC.41345“ (nicht-klassifizierter Kandidat nach NCBI)
- „P. sp. JD-2012“ (nicht-klassifizierter Kandidat nach NCBI)[10] – die bei Brisbin et al. (2018) in Fig. 2 angegebenen Zugriffsnummern für die Klade „Phaeo2“ endosymbiotischer Phaeocystis fallen in diese NCBI-Kandidaten-Spezies.[11]
Anmerkungen:
- Throndsen (1997) ordnet diese Gattung abweichend in die Ordnung Prymnesiales ein.[7]
- Die Spezies Phaeocystis giraudii (Derbès & Solier) De Toni, 1895 wurde in die Gattung Chrysoreinhardia verschoben als Synonym von Chrysoreinhardia giraudii (Derbès & Solier) C. Billard, 2000[7]
- Die Spezies Phaeocystis fuscescens (A. Braun} ex Kützing) De Toni 1895 wurde in die Gattung Tetrasporopsis verschoben als Synonym von Tetrasporopsis fuscescens (A. Braun ex Kützing) Lemmermann 1899[7]
- Weitere nicht-klassifizierte Kandidaten sind bei NCBI zu finden.[12]
- Zu den beiden endsymbiotischen Phaeocystis-Kladen „Phaeo1“ und „Phaeo2“ siehe unten.
Genom
BearbeitenEin Genomvergleich hat gezeigt, dass die RuBisCO-Spacer-Region (im Plastom, d. h. der Plastiden-DNA, zwischen zwei Untereinheiten des Enzyms Ribulose-1,5-bisphosphat-Carboxylase) unter eng verwandten kolonialen Phaeocystis-Arten hoch konserviert ist. Sie ist in P. antarctica, P. pouchetii und zwei warm-temperierten Stämmen von P. globosa identisch, es gibt lediglich eine einzige Basensubstitution in zwei kalt-temperierten Stämmen von P. globosa.[13]
Verbreitung und Lebenszyklus
BearbeitenPhaeocystis kann entweder in der Form freilebender Zellen oder als Kolonie existieren. Ein Sonderfall ist die endosymbiotische Lebensweise.
Fortpflanzung
BearbeitenEin haploid-diploider Lebenszyklus wurde bisher nur bei P. globosa beobachtet, wird aber auch bei anderen Arten wie P. pouchetii und P. antarctica vermutet. In diesem Zyklus ist die sexuelle Fortpflanzung bei der Bildung und dem Absterben der Kolonieblüte dominant. Abgesehen davon wurden zwei unterschiedliche Arten der vegetativen Fortpflanzung beobachtet.[14]
Freilebende Formen
BearbeitenFreilebende Formen von Phaeocystis sind weltweit verbreitet und kommen in einer Vielzahl von marinen Lebensräumen vor, darunter Küstenmeere, offene Ozeane, Polarmeere und im Meereis.[15]
Freilebende Zellen können je nach Art eine Vielzahl von Morphologien aufweisen. Alle Arten können als geschuppte Flagellaten (Geißeltierchen) existieren; bei P. scrobiculata und P. cordata ist dies sogar die einzige beobachtete Form.
Koloniebildende Formen und Algenblüten
BearbeitenDie Bildung schwimmender Kolonien ist eine besondere und kennzeichnende Eigenschaft der Gattung Phaeocystis. In einer solchen Kolonie sind Hunderte von Zellen in eine Matrix aus Polysaccharid-Gel eingebettet, die bei Algenblüten massiv an Größe zunehmen kann.[3]
Bei drei Arten wurden Kolonienbildung beobachtet (P. globosa, P. pouchetii und P. antarctica), diese können auch als ohne Schuppen und Filamente existieren.[14] Bei Kolonien von Phaeocystis kann eine Koloniehaut (en. colony skin) Schutz vor kleineren Zooplanktonfressern und Viren bieten.[16]
Die Allgegenwart und die Bildung großer Algenblüten machen die Gattung Phaeocystis zu einem wichtigen Bestandteil des Kohlenstoffkreislaufs im Ozean.[17][18] Außerdem produziert Phaeocystis Dimethylsulfid (DMS), einen Schlüsselakteur im Schwefelkreislauf.[19][20]
Die Bildung von Algenblüten in nährstoffreichen Gebieten wird durch die drei koloniebildenden Arten (P. globosa, P. pouchetii und P. antarctica) verursacht.[21] Diese können natürliche Ursache haben (z. B. im Rossmeer, in der Grönlandsee oder in der Barentssee)[22] oder durch anthropogene (menschenverursachte) Einträge hervorgerufen werden (z. B. in der Southern Bight in der südlichen Nordsee oder im Persischen Golf).
Im Allgemeinen blüht P. globosa in gemäßigten und tropischen Gewässern, während P. pouchetii und P. antarctica besser an die kalten Temperaturen angepasst sind, die in arktischen bzw. antarktischen Gewässern herrschen. P. pouchetii toleriert jedoch auch wärmere Temperaturen[23] und wurde tatsächlich schon in gemäßigten Gewässern gesehen.[24]
Die größten Phaeocystis-Blüten bilden sich in den Polarmeeren: P. pouchetii im Norden und P. antarctica im Süden.[1] Diese intensive Phaeocystis-Produktivität hält im Allgemeinen etwa drei Monate lang an und überspannt den größten Teil des Sommers auf der Südhalbkugel. Phaeocystis-reiche Ökosysteme stehen in der Regel mit kommerziell wichtigen Beständen von Krebstieren, Mollusken, Fischen und auch Säugetieren in Verbindung. Phaeocystis kann negative Auswirkungen auf höhere trophische Ebenen im marinen Ökosystem und damit auch auf menschliche Aktivitäten (z. B. Fischzucht und Küstentourismus) haben, indem sie während des Abklingens einer Blüte geruchsintensive Schäume an den Stränden bildet.[25]
Symbiose
Bearbeiten
Phaeocystis-Arten treten als Endosymbionten von Acantharien (Acantharea, informelle Gruppe der Strahlentierchen alias Radiolarien, wohl zu unterscheiden von der Pilzgattung Acantharia) auf.[26][11] Ob diese Symbiose einen echten Mutualismus darstellt, bei dem beide Partner profitieren, ist umstritten.[27] Aus verschiedenen Ozeanbecken gesammelte Acantharien beherbergen unterschiedliche Arten von Phaeocystis als dominante Symbionten:[26]
- P. antarctica wird als primärer Symbiont in Acanthariern im Südlichen Ozean (dem Antarktischen Ozean) gefunden.
- P. cordata und P. jahnii gehören zu den dominanten Symbionten in Akanthariern der warmen oligotrophen Regionen des Indischen und Pazifischen Ozeans.
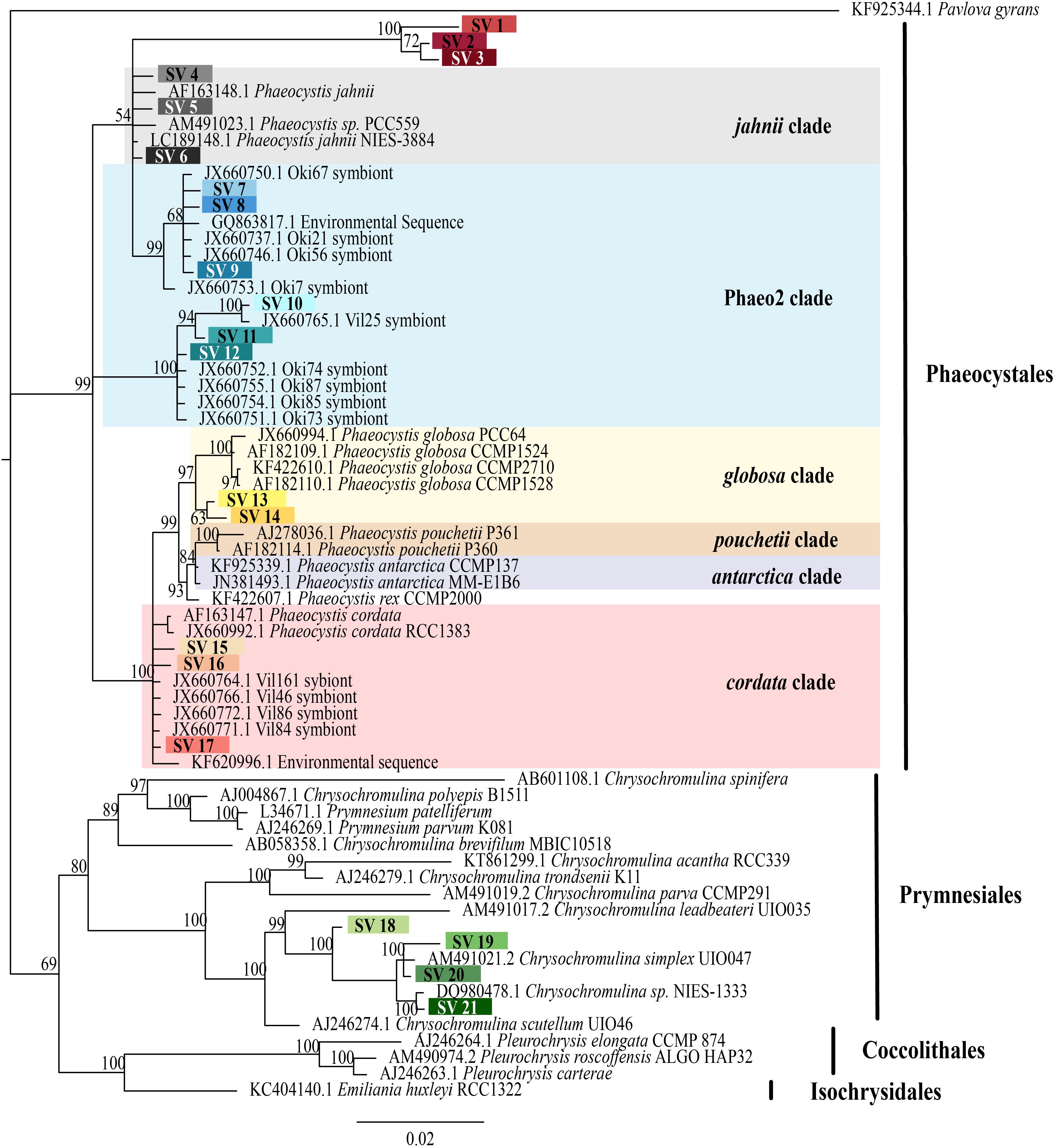
Zusätzlich zu den beschriebenen Phaeocystis-Arten machen Sequenzen aus einer mit „Phaeo2“ bezeichneten molekularen Klade oft einen Großteil der aus Acanthariern in warmen Wasserregionen gewonnenen symbiotischen Sequenzen aus (neben denen der sog. Klade „Phaeo1“[26][11][28], wohl zu unterscheiden von den gleichnamigen Kladen der Spezies Phaeodactylum tricornutum)[29][30]
Bei symbiotischen Phaeocystis-Arten wird ein extremer Zellumbau beobachtet, einschließlich einer drastischen Zunahme der Chloroplastenzahl und einer vergrößerten zentralen Vakuole.[26][11] Diese phänotypische Veränderung wird wahrscheinlich vom Wirt induziert, um die photosynthetische Leistung der Symbionten zu erhöhen. Die Endosymbiose wird jedoch zur Sackgasse für Phaeocystis, wenn sie die symbiotischen Zellen unfähig für weitere Zellteilungen macht.[27] Die Symbiose ist ökologisch relevant, weil sie Brennpunkte der Primärproduktion in nährstoffarmen Regionen schafft.[31] Es bleibt aber noch zu klären, inwieweit die Symbiose die Evolution von Phaeocystis beeinflusst hat.
Viren
BearbeitenVon den Arten P. globosa und P. pouchetii sind eine Reihe von Virusspezies mit Kandidatenstatus bekannt.[32] Diese wurden zunächst vorgeschlagen als Mitglieder der Virusgattung Prymnesiovirus, Familie Phycodnaviridae im Phylum Nucleocytoviricota (NCLDV). Nach neueren Untersuchungen sind einige der Kandidaten jedoch mit Gruppe der sog. „OLPG“ eher der Familie Mimiviridae (oder einer Schwesterfamilie Mesomimiviridae derselben) zuzuordnen.
Anmerkung: In der Prymnesiovirus-Schwestergattung Phaeovirus sind (trotz der Namensähnlichkeit) keine Phaeocystis-infizierende oder -assoziierte Viren bekannt.
- Familie Phycodnaviridae, Gattung Prymnesiovirus:[33]
- Spezies „Phaeocystis globosa virus“ (PgV)
- Spezies „Phaeocystis globosa virus 1“ (PgV-01T, T steht im Akronym für Texel, Niederlande)[34]
- Spezies „Phaeocystis globosa virus 2“ bis „11“ (PgV-02T bis PgV-11T)
- Spezies „Phaeocystis globosa virus 13“ (PgV-13T)
- Spezies „Phaeocystis globosa virus 15“ (PgV-15T)
- Spezies „Phaeocystis globosa virus 17“ (PgV-17T)
- Spezies „Phaeocystis globosa virus 18“ (PgV-18T)
- Spezies „Phaeocystis globosa virus 102“ (PgV-102P, P steht für Plymouth, Südwestengland)[35]
- Verschiebe-Option Richtung Familie Mimiviridae:
- Mit Phaeocystis assoziierte Virophagen (Klasse Maveriviricetes)
Man vermutet, dass die Algenblühen verursachende Mikroorganismen durch sie parasitierende Viren kontrolliert werden und diese sogar in der Lage sind, eine Blüte zusammenbrechen zu lassen.
Globale Auswirkungen
BearbeitenDie Gattung Phaeocystis ist ein wichtiger Produzent von 3-Dimethylsulphoniopropionat (DMSP). DMSP wird von weiteren marinen Mikroben in flüchtige Schwefelverbindungen zersetzt, insbesondere Methanthiol (CH3SH) und Dimethylsulfid (DMS, (CH3)2S).[1] Ein Großteil des DMS im Meerwasser wird zu Dimethylsulfoxid (DMSO, (CH3)2S=O) oxidiert.[45] Biogenes DMS trägt jährlich etwa 1.5×1010 kg Schwefel zur Atmosphäre bei und spielt daher eine wichtige Rolle im globalen Schwefelkreislauf, der die Wolkenbildung und möglicherweise die Klimaregulierung beeinflussen kann.[1]
Einzelnachweise
Bearbeiten- ↑ a b c d Phaeocystis Research. Abgerufen am 28. Oktober 2015.
- ↑ a b Véronique Schoemann, Sylvie Becquevort, Jacqueline Stefels, Véronique Rousseau, Christiane Lancelot: Phaeocystis blooms in the global ocean and their controlling mechanisms: a review. In: Journal of Sea Research, Serie: Iron Resources and Oceanic Nutrients - Advancement of Global Environmental Simulations. 53. Jahrgang, Nr. 1–2, 1. Januar 2005, S. 43–66, doi:10.1016/j.seares.2004.01.008, bibcode:2005JSR....53...43S.
- ↑ a b Welcome to the Phaeocystis antarctica genome sequencing project homepage. Abgerufen am 28. Oktober 2015.
- ↑ Sara J. Bender, Dawn M. Moran, Matthew R. McIlvin, Hong Zheng, John P. McCrow, Jonathan Badger, Giacomo R. DiTullio, Andrew E. Allen, and Mak A. Saito: Colony formation in Phaeocystis antarctica: connecting molecular mechanisms with iron biogeochemistry, in: Biogeosciences, Band 15, Nr. 16, 21. August 2018, S. 4923–4942, doi:10.5194/bg-15-4923-2018
- ↑ Robert A. Andersen, J. Craig Bailey, Johan Decelle, Ian Probert: Phaeocystis rex sp. nov. (Phaeocystales, Prymnesiophyceae): a new solitary species that produces a multilayered scale cell covering. In: European Journal of Phycology. 50. Jahrgang, Nr. 2, 3. April 2015, ISSN 0967-0262, S. 207–222, doi:10.1080/09670262.2015.1024287 (englisch).
- ↑ WoRMS: Phaeocystis Lagerheim, 1893
- ↑ a b c d AlgaeBase: Phaeocystis Lagerheim, 1893
- ↑ NCBI: Phaeocystis (genus); graphisch: Phaeocystis, auf: Lifemap, NCBI Version.
- ↑ Robert A. Andersen, J. Craig Bailey, Johan Decelle, Ian Probert: Phaeocystis rex sp. nov. (Phaeocystales, Prymnesiophyceae): a new solitary species that produces a multilayered scale cell covering, in: European Journal of Phycology, Band 50, Nr. 2, April 2015, doi:10.1080/09670262.2015.1024287
- ↑ NCBI: Phaeocystis sp. JD-2012 (species)
- ↑ a b c d Margaret Mars Brisbin, Mary M. Grossmann, Lisa Y. Mesrop, Satoshi Mitarai: Intra-host Symbiont Diversity and Extended Symbiont Maintenance in Photosymbiotic Acantharea (Clade F). In: Frontiers in Microbiology. 9. Jahrgang, 27. August 2018, ISSN 1664-302X, S. 1998, doi:10.3389/fmicb.2018.01998, PMID 30210473, PMC 6120437 (freier Volltext) – (englisch). „Phaeo2“ siehe Fig. 2
- ↑ NCBI: unclassified Phaeocystis (list)
- ↑ Martin Lange, Yue-Qin Chen, Linda K. Medlin: Molecular genetic delineation of Phaeocystis species (Prymnesiophyceae) using coding and non-coding regions of nuclear and plastid genomes. In: European Journal of Phycology. 37. Jahrgang, Nr. 1, 1. Februar 2002, ISSN 1469-4433, S. 77–92, doi:10.1017/S0967026201003481 (awi.de [PDF]).
- ↑ a b Véronique Rousseau, Marie-Josèphe Chrétiennot-Dinet, Anita Jacobsen, Peter Verity, Stuart Whipple: The life cycle of Phaeocystis: state of knowledge and presumptive role in ecology. In: Biogeochemistry. 83. Jahrgang, Nr. 1–3, 13. April 2007, ISSN 0168-2563, S. 29–47, doi:10.1007/s10533-007-9085-3.
- ↑ Helge Abildhauge Thomsen, Kurt. R. Buck, F. P. Chavez: Haptophytes as components of marine phytoplankton, in: J. C. Green, B. S. C. (Hrsg.): The Haptophyte Algae, Clarendon Press, Oxford, UK, Januar 1994, S. 187–208, ResearchGate
- ↑ Peter G. Verity, Corina P. Brussaard, Jens C. Nejstgaard, Maria A. van Leeuwe, Christiane Lancelot, Linda K. Medlin: Current understanding of Phaeocystis ecology and biogeochemistry, and perspectives for future research. In: Biogeochemistry. 83. Jahrgang, Nr. 1–3, 16. März 2007, ISSN 0168-2563, S. 311–330, doi:10.1007/s10533-007-9090-6 (awi.de [PDF]).
- ↑ Walker O. Smith, Louis A. Codispoti, David M. Nelson, Thomas Manley, Edward J. Buskey, H. Joseph Niebauer, Glenn F. Cota: Importance of Phaeocystis blooms in the high-latitude ocean carbon cycle. In: Nature. 352. Jahrgang, Nr. 6335, 8. August 1991, S. 514–516, doi:10.1038/352514a0, bibcode:1991Natur.352..514S.
- ↑ G. R. DiTullio, J. M. Grebmeier, K. R. Arrigo, M. P. Lizotte, D. H. Robinson, A. Leventer, J. P. Barry, M. L. VanWoert, R. B. Dunbar: Rapid and early export of Phaeocystis antarctica blooms in the Ross Sea, Antarctica. In: Nature. 404. Jahrgang, Nr. 6778, 6. April 2000, S. 595–598, doi:10.1038/35007061, PMID 10766240 (nature.com).
- ↑ J. Stefels, W. H. M. Van Boekel: Production of DMS from dissolved DMSP in axenic cultures of the marine phytoplankton species Phaeocystis sp., in: Mar. Ecol. Prog. Ser., Band 97, 8. Juli 1993, S. 11–18.
- ↑ J. Stefels, L. Dijkhuizen, W. W. C. Gieskes: DMSP-lyase activity in a spring phytoplankton bloom off the Dutch coast, related to Phaeocystis sp. abundance. In: Marine Ecology Progress Series. 123. Jahrgang, 20. Juli 1995, S. 235–243, doi:10.3354/meps123235, bibcode:1995MEPS..123..235S (rug.nl [PDF]).
- ↑ Christiane Lancelot, M. D. Keller, Véronique Rousseau, Walker O. Smith Jr, Sylvie Mathot: Autecology of the marine haptophyte Phaeocystis sp. (sic!), in: David M. Anderson, Allan D. Cembella, Gustaaf M. Hallegraeff (Hrsg.): Physiological ecology of harmful algal blooms, NATO ASI Series G: Ecological Sciences, Band 41, Springer-Verlag, Berlin, 2. Juni 1998, S. 209–224, ISBN 978-3540641179 (Buch: freier Volltext) – Der Titel des Artikels wird oft als ‚Autoecology‘ verschrieben.
- ↑ A. Orkney, T. Platt, B. E. Narayanaswamy, I. Kostakis, H. A. Bouman: Bio-optical evidence for increasing Phaeocystis dominance in the Barents Sea, in: Philosophical Transactions of the Royal Society A: Mathematical, Physical and Engineering Sciences, 31. August 2020, doi:10.1098/rsta.2019.0357
- ↑ M. E. M. Baumann, C. Lancelot, F. P. Brandini, E. Sakshaug, D. M. John: The taxonomic identity of the cosmopolitan prymnesiophyte Phaeocystis: a morphological and ecophysiological approach. In: Journal of Marine Systems. 5. Jahrgang, Nr. 1, 1994, S. 5–22, doi:10.1016/0924-7963(94)90013-2, bibcode:1994JMS.....5....5B.
- ↑ Catharina J. M. Philippart, Gerhard C. Cadée, Wim van Raaphorst, Roel Riegman: Long-term phytoplankton-nutrient interactions in a shallow coastal sea: Algal community structure, nutrient budgets, and denitrification potential. In: Limnology and Oceanography. 45. Jahrgang, Nr. 1, 1. Januar 2000, ISSN 1939-5590, S. 131–144, doi:10.4319/lo.2000.45.1.0131, bibcode:2000LimOc..45..131P.
- ↑ C. Lancelot, S. Mathot: Dynamics of a Phaeocystis-dominated spring bloom in Belgian coastal waters. I. Phytoplanktonic activities and related parameters. In: Marine Ecology Progress Series. 37. Jahrgang, 1987, S. 239–248, doi:10.3354/meps037239, bibcode:1987MEPS...37..239L.
- ↑ a b c d M. Johan Decelle, Rafel Simó, Martí Galí, Colomban de Vargas, Sébastien Colin, Yves Desdevises, Lucie Bittner, Ian Probert, Fabrice Not: An original mode of symbiosis in open ocean plankton. In: Proceedings of the National Academy of Sciences. 109. Jahrgang, Nr. 44, 30. Oktober 2012, ISSN 0027-8424, S. 18000–18005, doi:10.1073/pnas.1212303109, PMID 23071304, PMC 3497740 (freier Volltext), bibcode:2012PNAS..10918000D (englisch). Zu „Phaeo1“ und „Phaeo2“ siehe Fig. 1, zu „Phaeo2“ auch Fig. 3.
- ↑ a b Johan Decelle: New perspectives on the functioning and evolution of photosymbiosis in plankton. In: Communicative & Integrative Biology. 6. Jahrgang, Nr. 4, 30. Juli 2013, S. e24560, doi:10.4161/cib.24560, PMID 23986805, PMC 3742057 (freier Volltext).
- ↑ M. Johan Decelle: Ecologie et évolution de la photosymbiose chez les acanthaires (radiolaires), Dissertation an der Universität Pierre und Marie Curie (UPMC), Paris, 11. Januar 2018 (französisch)
- ↑ A Look Back at the U.S. Department of Energy’s Aquatic Species Program: Biodiesel from Algae, National Renewable Energy Laboratory (NREL/TP-580-24190) – Gegenbeispiel
- ↑ NCBI: Phaeodactylum (genus, Bacillariophyceae)
- ↑ David A. Caron, Neil R. Swanberg, Anthony F. Michaels, Frances A. Howse: Primary productivity by symbiont-bearing planktonic sarcodines (Acantharia, Radiolaria, Foraminifera) in surface waters near Bermuda. In: Journal of Plankton Research. 17. Jahrgang, Nr. 1, 1. Januar 1995, ISSN 0142-7873, S. 103–129, doi:10.1093/plankt/17.1.103 (englisch).
- ↑ a b c Romain Blanc-Mathieu, Håkon Dahle, Antje Hofgaard, David Brandt, Hiroki Ban, Jörn Kalinowski, Hiroyuki Ogata, Ruth-Anne Sandaa: A Persistent Giant Algal Virus, with a Unique Morphology, Encodes an Unprecedented Number of Genes Involved in Energy Metabolism ( des vom 21. Juli 2021 im Internet Archive) Info: Der Archivlink wurde automatisch eingesetzt und noch nicht geprüft. Bitte prüfe Original- und Archivlink gemäß Anleitung und entferne dann diesen Hinweis., in: ASM Journals: Journal of Virology, Band 95, Nr. 8, 25. März 2021, doi:10.1128/JVI.02446-20 .
PrePrint: bioRxiv, doi:10.1101/2020.07.30.228163, CSH, 13. Januar 2021 - ↑ NCBI: Prymnesiovirus (Genus)
- ↑ a b Jean-Michel Claverie, Chantal Abergel: Mimiviridae: An Expanding Family of Highly Diverse Large dsDNA Viruses Infecting a Wide Phylogenetic Range of Aquatic Eukaryotes]. In: Viruses. 2018 Sep; 10(9), 18. September 2018, S. 506, doi:10.3390/v10090506, PMC 6163669 (freier Volltext), PMID 30231528, Tab. 2
- ↑ Willie Wilson, Declan C. Schroeder, Jenna Ho, Martin Canty: Phylogenetic analysis of PgV-102P, a new virus from the English Channel that infects Phaeocystis globosa, in: J Mar Biol Ass UK 86, Juni 2006, S. 485–490, doi:10.1017/S0025315406013385
- ↑ a b c Jônatas Abrahão, Lorena Silva, Ludmila Santos Silva, Jacques Yaacoub Bou Khalil, Rodrigo Rodrigues, Thalita Arantes, Felipe Assis, Paulo Boratto, Miguel Andrade, Erna Geessien Kroon, Bergmann Ribeiro, Ivan Bergier, Herve Seligmann, Eric Ghigo, Philippe Colson, Anthony Levasseur, Guido Kroemer, Didier Raoult, Bernard La Scola: Tailed giant Tupanvirus possesses the most complete translational apparatus of the known virosphere. In: Nature Communications. 9. Jahrgang, Nr. 1, 27. Februar 2018, doi:10.1038/s41467-018-03168-1 (nature.com).
- ↑ Sebastien Santini, Sandra Jeudy, Julia Bartoli, Olivier Poirot, Magali Lescot, Chantal Abergel, Valérie Barbe, K. Eric Wommack, Anna A. M. Noordeloos, Corina P. D. Brussaard, Jean-Michel Claverie: Genome of Phaeocystis globosa virus PgV-16T highlights the common ancestry of the largest known DNA viruses infecting eukaryotes, in: Proc Natl Acad Sci USA 110(26), 25. Juni 2013, S. 10800–10805, doi:10.1073/pnas.1303251110, PMC 3696832 (freier Volltext), PMID 23754393
- ↑
Yanze Li, Hisashi Endo, Yasuhiro Gotoh, Hiroyasu Watai, Nana Ogawa, Romain Blanc-Mathieu, Takashi Yoshida, Hiroyuki Ogata: The Earth Is Small for “Leviathans”: Long Distance Dispersal of Giant Viruses across Aquatic Environments, in: Microbes Environ. 34(3), September 2019, S. 334–339, doi:10.1264/jsme2.ME19037, PMC 6759346 (freier Volltext), PMID 31378760; insbes. Fig. 4.
Anm.: Wegen der Platzierung im Stammbaum und DORTIGER Referenz auf Santini (2013) meint Phaeocystis_globosa_virus hier Tethysvirus hollandense, insbesondere PgV-16T. Mit ‘Megaviridae’ werden die erweiterten Mimiviridae, also die Imitervirales bezeichnet, mit ‘Megamimivirinae’ die ganze Familie Mimiviridae und ‚Mesomimivirinae‘ umfasst alle drei Schwesterfamilien der Mimiviridae (Allo-, Meso- und Schizomimivirinae). - ↑ a b Andrew M. Q. King, Michael J. Adams et al.: Virus Taxonomy. Elsevier, San Diego 2012, ISBN 978-0-12-384684-6, Family - Phycodnaviridae, S. 249–262, doi:10.1016/B978-0-12-384684-6.00024-0.
- ↑ NCBI: Phaeocystis pouchetii virus (species)
- ↑ Lucie Gallot-Lavallee, Guillaume Blanc, Jean-Michel Claverie: Comparative genomics of Chrysochromulina Ericina Virus (CeV) and other microalgae-infecting large DNA viruses highlight their intricate evolutionary relationship with the established Mimiviridae family ( des vom 18. Oktober 2019 im Internet Archive) Info: Der Archivlink wurde automatisch eingesetzt und noch nicht geprüft. Bitte prüfe Original- und Archivlink gemäß Anleitung und entferne dann diesen Hinweis., in: J. Virol., 26 April 2017, doi:10.1128/JVI.00230-17
- ↑ Said Mougari, Dehia Sahmi-Bounsiar, Anthony Levasseur, Philippe Colson, Bernard La Scola: Virophages of Giant Viruses: An Update at Eleven. In: Viruses. Band 11, Nr. 8, 2019, ISSN 1999-4915, S. 733, doi:10.3390/v11080733, PMID 31398856, PMC 6723459 (freier Volltext).
- ↑ Meriem Bekliz, Philippe Colson, Bernard La Scola: The Expanding Family of Virophages. In: MDPI Viruses. Band 8, Nr. 11 (Special Issue Viruses of Protozoa), 23. November 2016, S. 317, doi:10.3390/v8110317.
- ↑ NCBI: Phaeocystis globosa virus virophage. (species) Auf: ncbi.nlm.nih.gov; zuletzt abgerufen am 22. März 2021.
- ↑ Martí Galí, Rafel Simó: Occurrence and cycling of dimethylated sulfur compounds in the Arctic during summer receding of the ice edge ( des vom 24. Februar 2016 im Internet Archive) Info: Der Archivlink wurde automatisch eingesetzt und noch nicht geprüft. Bitte prüfe Original- und Archivlink gemäß Anleitung und entferne dann diesen Hinweis. (PDF), in: Marine Chemistry, Band 122, Ausgabe 1–4, Oktober 2010, S. 105–117, doi:10.1016/j.marchem.2010.07.003, Epub 24. Juli 2010.
{kind=link}
{kind=link}
{kind=link}