Cruciviren
Als Cruciviren (englisch Cruciviruses, CruVs) werden gewöhnlich im engeren Sinne solche chimäre Viren verstanden,
- das ein Replikations-Initiator-Protein für Rolling-Circle-Replikation (als REP, Rep, genauer RC-Rep bezeichnet) kodiert und
- aufgrund seiner charakteristischen konservierte Haarnadelstruktur (en. stem-loop structure) mit einer Nonanukleotid-Sequenz,[Anm. 1] die als Replikationsursprung (Ori) dient, sie als Mitglieder des Phylums Cressdnaviricota (en. Circular Rep-encoding ssDNA viruses, CRESS-DNA-Viren oder CressDNA-Viren) ausweist,
- deren Gen für das Kapsidprotein (als CAP, Cp, CP oder Cp bezeichnet) aber Homologie zu Kapsidgenen von RNA-Viren zeigt (und nicht zu anderen herkömmlichen CRESS-DNA-Viren-Viren).[1]
Der Begriff wird daher üblicherweise synonym zu RNA-DNA-Hybrid-Virus (RDHV) verwendet.[Anm. 2]
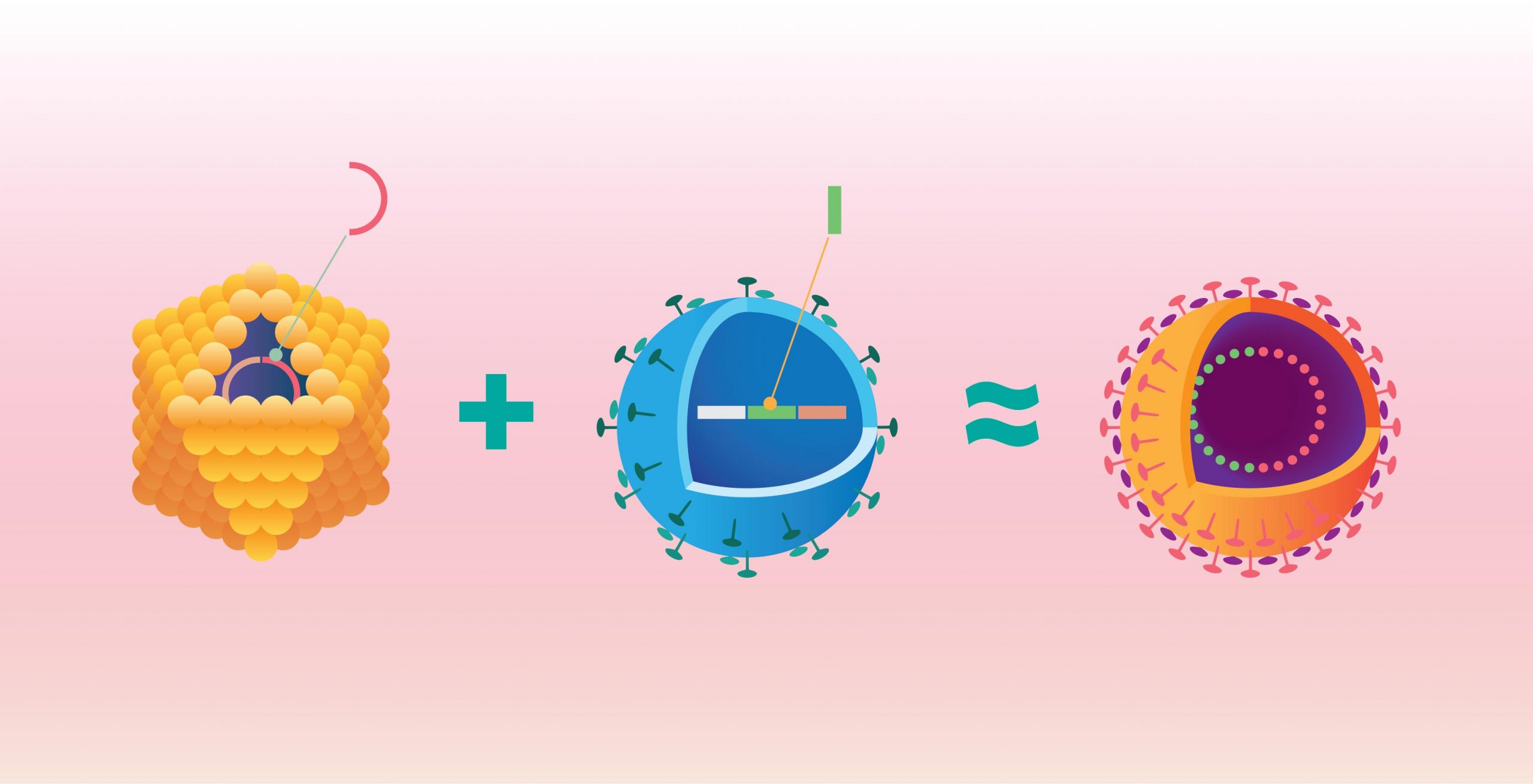
Die Viruspartikel (Virionen) der Cruciviren ähneln also aufgrund ihrer durch das Kapsidprotein bestimmten äußeren Gestalt und Größe (Morphologie) bestimmten RNA-Viren, die innere Genom-Architektur stimmt dagegen mit anderen (üblicherweise viel kleineren) CressDNA-Viren überein.
Der erste Vertreter der Cruciviren ist das „Boiling Springs Lake RNA-DNA Hybrid Virus“ (BSL-RDHV, auch BSL_RDHV) aus einer 2012 veröffentlichten Studie, ein weiterer ist „Idotea-Virus IWaV278“ (Idotea virus IWaV278, „Idotea wosenenskii associated virus“, IWaV-278).
Die entscheidende Frage ist, ob diese insgesamt (oder wenigstens zu einem überwiegenden Teil) aus einem einzelnen (erd-)geschichtlichen RNA-DNA-Hybridisierungsereignis hervorgehen, und daher (bis auf „Ausnahmen“) eine monophyletische Verwandtschaftsgruppe (Klade) bilden. Für diesen Fall wurde für die BSL-RDHV-ähnlichen Viren (en. „BSL-RDHV-like viruses“) die taxonomische Rangstufe einer Familie mit Namen „Cruciviridae“ vorgeschlagen. Das Thema ist aber aufgrund der hohen Diversität speziell in dieser Gruppe, aber auch unter den CressDNA-Viren allgemein, äußerst komplex und derzeit (Stand März 2021) immer noch in der Diskussion. Dieser Artikel soll diese Entwicklung in groben Zügen nachzeichnen.
Entdeckung
BearbeitenSeit 2001 wurden von Kenneth M. Stedman und Kollegen in den Geothermalgebieten im Lassen-Volcanic-Nationalpark (Geothermal areas in Lassen Volcanic National Park, englisch) in Nordkalifornien eine Reihe von Untersuchungen, darunter insbesondere Metagenomanalysen durchgeführt.[2] Dabei wurde im dortigen Boiling Springs Lake (40,4355° N, 121,3971° W)[3][2] eine Gensequenz (Contig/MAG) mit einem Aufbau wie bei den CRESS-DNA-Viren (dem heutigen Phylum Cressdnaviricota) gefunden. Insbesondere zeigte das Replikations-Initiations-Protein Rep Homologien zum Porcinen Circovirus-2 (PCV2, Gattung Circovirus). Das Kapsidprotein CAP war jedoch homolog zu dem von Viren der Familie Tombusviridae.[2][1]
Diese chimären (oder hybriden) Genomsequenz wurde vorschlagsgemäß einem neuen Virus mit der Bezeichnung „Boiling Springs Lake RNA-DNA Hybrid Virus“ (BSL-RDHV, auch BSL_RDHV) zugeordnet (Diemer und Stedman, 2012).[4][2][1] Aufgrund der Analysen wurden als mögliche BSL-RDHV-Wirte Mitglieder der Supergruppe SAR (Stramenopiles/Alveolata/Rhizaria) vorgeschlagen.[5]
Weitere Funde und Kandidaten
BearbeitenSeit dem Fund von BSL-RDHV wurden in weiteren Metagenomanalysen aus einer Vielzahl von Umgebungen etliche weitere chimäre Virus-Sequenzen als Kandidaten die Gruppe der Criciviren gefunden. Beispiele für solche Habitate sind die Tampa-Bucht, die Finger Lakes (im Norden des US-Bundesstaates New York), Seen der Antarktis, Tiefseesedimente und im Inneren von Libellen (Arvind Varsani, Mya Breitbart).[2][3][6][7][8]
Einige Beispiele:[Anm. 3]
- „Boiling Springs Lake RNA-DNA Hybrid Virus“ (BSL-RDHV, BSL_RDHV)[2][1][9][2][4][Anm. 4]
- „Idotea-Virus IWaV278“ (Idotea virus IWaV278, „Idotea wosenenskii associated virus“, IWaV-278)[10] – gefunden in Asseln der Gattung Idotea an der US-Pazifikküste bei Port Townsend, Washington (Bundesstaat)[11][12][8]
- „RDHV-like virus SF1“ – Fundort: San Francisco, Abwasser[13][8]
- „Fire ant associated circular virus 1“ (FaACV-1)[14][1][15][Anm. 5]
- „Faecal-associated circular virus-13“ (FaCV-13) alias „Sewage-associated circular virus-13“ (SaCV-13)[16][17]
- „Chimeric virus 1“ (CHIV1) bis „Chimeric virus 13“ (CHIV13)[1]
- „Chimeric virus 14“ (CHIV14)[18][Anm. 6][8][19]
- „Peat Bog 1 RNA-DNA hybrid virus“ (PB1-RDHV) – Fundort: „Peat Bog“ marsh, Woodburn, Oregon, USA.[2][Anm. 7][Anm. 8]
- „Mill Creek crucivirus 1“ (CruV-MC1), Fundort: Torfmoor am Mill Creek (45,1539° N, 122,8445° W), Woodburn, Oregon, USA.[Anm. 8]
- „Mill Creek crucivirus 2“ (CruV-MC2) – Fundort: Torfmoor am Mill Creek, Woodburn
- „Mill Creek crucivirus 3“ (CruV-MC3) – Fundort: im Wasser/Bachbett vom Mill Creek, Woodburn[20]
- „Crucivirus-15“ (CruV-15-B) bis „Crucivirus-51“ (CruV-51-F)[17][8][Anm. 9]
- „Crucivirus-81“ (CruV-81) bis „Crucivirus-540“ (CruV-540) – nicht alle Nummern besetzt.[5][21][Anm. 5]
- „Crucivirus-like circular genetic element-85“ (CruV-85)[5][22]
- „Crucivirus-like circular genetic element-471“ (CruV-471)[5][23]
Andere RNA-DNA-Hybridisierungsereignisse
BearbeitenDas für die Entstehung der Cruciviren als RNA-DNA-Hybridviren vorausgesetzte Hybridisierungsereignis setzt (das als Resultat wieder ein DAN-Virus entsteht) eine „reverse“ Transkription von RNA zu DNA voraus. Allerdings besitzen die gefundenen Cruciviren im Gegensatz zu den Retroviren nicht selbst ein dafür nötiges Enzym (d. h. eine Reverse Transkriptase). Man hatte daher zunächst angenommen, dass ein derartiges Hybridisierungsereignis in der Evolutionsgeschichte der (CRESS-)DNA-Viren einmalig ist.
Weitere Studien belegten aber offenbar, dass es in der Evolution doch mehrere solche RNA-DNA-Hybridisierungsereignissse, insbesondere auch im Phylum Cressdnaviricota, so dass nicht alle RNA-DNA-Hybrid-Viren per se einer einzigen Verwandtschaftsgruppe (Klade) angehören.
DfCyclV
BearbeitenEin vermutliches Beispiel dafür, dass nicht bei alle RNA-DNA-Hybridviren das CP von den Tombusviridae stammt, zeigte sich bei Untersuchung der Vielfalt von mit Libellen assoziierten ssDNA-Viren. Ein dabei (neben anderen) gefundenes Virus-Genom mit der Bezeichnung „Dragonfly cyclisvirus“[1] – offenbar ist gemeint „Dragonfly cyclicusvirus“ (DfCyclV),[24][25] Spezies Strenuaivirus antaflavis – zeigte eine schwache, aber signifikante Homologie eines mutmaßlichen Proteins zum CP des Tabak-Mosaik-Satelliten-Virus (alias Satelliten-Tabaknekrosevirus, en. Satellite tobacco necrosis virus, STMV)[26] der Spezies Tobacco virtovirus 1 (Gattung Virtovirus).[27][25] Das CP des Satelliten-Tabaknekrosevirus unterscheidet sich jedoch in Sequenz und Struktur grundlegend von denen der Tombusviridae und ähnelt am ehesten den CPs der Geminiviridae. Daher ist vermutlich DfCyclV ein CRESS-DNA-Virus, aber kein Mitglied der Klade der BSL-RDHV-ähnlichen Viren.[1]
SsHADV-1
BearbeitenÜber einen Fund von RDHVs anderen Typs berichteten zudem Koonin und Dolja (2012): Sclerotinia sclerotiorum hypovirulence-associated DNA virus 1 (SsHADV-1), ein Geminiviridae-ähnliches Mykovirus mit zyklischer ssDNA, aber mit signifikanten Unterschieden in Genom-Organisation und Morphologie.[28][29] Auch für die Bacilladnaviridae wird eine ähnliche evolutionäre Vergangenheit angenommen.[30]
Bidnaviridae
BearbeitenZudem berichteten Krupovic und Koonin (2014), dass eine andere ssDNA-Familie, die Bidnaviridae, ebenfalls eine „turbulente“ evolutionäre Vergangenheit hat, u. a. mit einer mutmaßlichen Gen-Übernahme von den dsRNA-Viren der Reoviridae.[31]
Bacilladnaviridae
BearbeitenDarüber hinaus wurde die Übernahme von Kapsiden mit einer T=3-Symmetrie aus der ssRNA-Familien Nodaviridae durch CRESS-Viren als Ursprung der Bacilladnaviridae. Die Nodaviridae sind wie die Tombusviridae ein Mitglied der Kitrinoviricota (Orthornavirae), die Bacilladnaviridae wie die Cruciviren Mitglied der Cressdnaviricota.[32]
Weitere
BearbeitenTisza et al. präsentierten 2020 Belege für zusätzliche unabhängige Rekombinationsereignisse zwischen CRESS-Viren und ssRNA-Viren sowie ssDNA-Bakteriophagen.[32]
REP-Polyphylie
BearbeitenUm von einem einzigen RNA-DNA-Hybridisierungsereignis ausgehen zu können, ist es daher notwendig, in der Gruppe der Cruciviren Homologie (Biologie) des (von RNA-Viren stammenden) CAP-Gens vorauszusetzen. Überraschenderweise besteht aber auch bei Beschränkung auf BSL-RDHV-ähnliche CAP-Sequenzen beim REP-Protein offenbar keine Monophylie. Weitere phylogenetische Untersuchungen der Virensequenzen chimärer Virusgenome mit Bezeichnung „Chimeric virus1“ (CHIV1) bis „Chimeric virus13“ (CHIV13) aus verschiedenen Umgebungen durch Simon Roux et al. (2013)[1] zeigten nur zum Teil Homologie des Kapsidproteins CAP zu den ssRNA-Viren der Tombusviridae. Zum Teil gab es aber Homologie zu Sequenzen der den bisher – Stand 21. Juni 2021 – nicht klassifizierten Spezies „Sclerophthora macrospora virus A“ (SmV-A)[33] und „Plasmopara halstedii virus A“ (PhV-A)[34] – beide sind assoziiert[Anm. 10] mit Eipilzen[Anm. 11] der Familie Peronosporaceae.[1]
Diese mosaikartige Zusammensetzung des Genoms in der Gruppe der Cruciviren erwies sich als das Hauptproblem ihrer taxonomischen Klassifizierung.
Möglicher nachträglicher Austausch von REP
BearbeitenRoux et al. (2013) versuchten die angenommene Monophylie der Cruciviren durch folgende Annahme aufrechtzuerhalten:
Der Transfer eines CAP-Gens von RNA-Viren auf DNA-Viren (genauer CressDNA-Viren) sollte ein einmaliges (oder sehr seltenes) Ereignis sein. Der Erwerb eines solchen Gens von der Familie Tombusviridae könnte dann der Ursprung einer Klade (Verwandtschaftsgruppe) von BSL-RDHV-ähnlichen Viren sein. Das bei der Hybridisierung von den Tombusviridae übernommene CAP bot offenbar mehr Platz, was einen evolutionären Vorteil darstellt. Die so entstandenen Hybridviren hatten im Kapsid mehr Platz und konnten in der Folge auch ihr Genom (weiter) vergrößern.[1] Dazu passt, dass die von Roux et al. 2013 untersuchten Genome der Cruciviren im Vergleich zu herkömmlichen CRESS-DNA-Viren eine große Länge zeigten.
Die Vielfalt der REP-Gene bei den untersuchten Sequenzen wird als nachträglicher als Austausch dieses Gens durch das verschiedener anderer CRESS-DNA-Viren gedeutet. Man hat spekuliert, dass direkt nach der Hybridisierung, d. h. der Übernahme des Gens für CAP, dieses mit dem Gen für das REP noch nicht ausreichend kompatibel war. Im Rahmen weiterer Anpassungen könnte es dann per horizontalem Gentransfer (HGT) zu einer Ersetzung des REP durch das von verwandten Viren, d. h. CRESS-DNA-Viren, gekommen sein. Dies wäre wohl viel leichter möglich und daher wahrscheinlicher ist als die ursprüngliche Übernahme des Gens für CAP von RNA-Viren, so dass ein solcher Vorgang wiederholt stattgefunden haben könnte.[1]
Vorschlag einer Familie Cruciviridae
Bearbeiten| „Cruciviridae“ | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Systematik | ||||||||||||||
| ||||||||||||||
| Taxonomische Merkmale | ||||||||||||||
| ||||||||||||||
| Wissenschaftlicher Name | ||||||||||||||
| „Cruciviridae“ | ||||||||||||||
| Links | ||||||||||||||
|
Die Überlegungen führten schließlich dazu, für die BSL-RDHV-ähnlichen chimären Viren (BSL-RDHV-like viruses) innerhalb des Phylums Cressdnaviricota die Bezeichnung „Cruciviren“ (en. cruciviruses, CruVs) einzuführen. Insgesamt waren von diesen mit Stand Oktober 2020 nicht weniger als 461 Kandidaten bekannt.[2][3]
Darüber hinaus wurde unter der Annahme, dass diese eine Verwandtschaftsgruppe (Klade) bilden, für diese provisorisch der taxonomische Rang einer Familie „Cruciviridae“ innerhalb dieses Phylums vorgeschlagen.[2][36]
Klassifizierung (Versuch)
BearbeitenRoux et al (2013) haben eine Klassifizierung der Cruciviren – der postulierten Familie „Cruciviridae“ – nach Homologie des REP zu verschiedenen Familien der Cressdnaviricota vorgeschlagen (siehe dort insbes. Fig. 1 und Suppl. 2)[1] Ungeklärt blieb aber, ob diese Gruppen aber angesichts der leichten Ersetzbarkeit des REP monophyletisch sind.[Anm. 12][1]
Familie(?): „Cruciviridae“ (Taxonomie, bzw. Klassifizierung der Cruciviren nach REP)[1][17][8]
- Klade(?): Circo-like REP-associated gene – REP-Homologie zum Porcinen Circovirus-2 (PCV-2, Circoviridae)
- BSL-RDHV; CHIV1, CHIV2, CHIV3, CHIV4, CHIV5; RDHV-SF1 (Krupovic), CRUV-15-B, CRUV-19-B, CRUV-20-B, CRUV-21-B, CRUV-22-F, CRUV-23-F, CRUV-24-F, CRUV-25-B, CRUV-32-F, CRUV-33-F, CRUV-34-B, CRUV-35-B, CRUV-36-B, CRUV-38-B, CRUV-39-B, CRUV-40-B, CRUV-41-F, CRUV-42-B, CRUV-43-F, CRUV-44-B, CRUV-45-F, CRUV-46-B, CRUV-47-F (Quaiser)
- IWaV278 (Bistolas)
- Klade(?): Nano-like Rep-associated gene – REP-Homologie zum Milk vetch dwarf virus (MDV, Nanoviridae)
- CHIV6, CHIV7, CHIV8, CHIV9, CHIV10, CHIV11, CHIV12; CRUV-16-B, CRUV-18-B, CRUV-26-F, CRUV-27-B, CRUV-28-F, CRUV-29-F, CRUV-31-B, CRUV-48-B CRUV-49-B, CRUV-50-F, CRUV-51-F (Roux, Quaiser)
- Klade(?): Gemini-like Rep-associated gene – REP-Homologie zum Maize Streak Virus (MSV, Geminiviridae)
- CHIV13, CHIV14; CRUV-17-B, CRUV-30-B, CRUV-37-F, FaCV-13 (Quaiser)
Die CHIV-Genome zeigten die für Cressdnaviricota charakteristischen Haarnadelstruktur (en. stem loop) mit einer Nonanukleotid-Sequenz, die bei diesen Viren als Replikationsursprung dienen. Neben den CAP- und REP-Genen enthalten einige der CHIVs laut Vorhersage bis zu vier zusätzliche Offene Leserahmen (en. open reading frames, ORFs).[1]
Die Analyse des Genoms von IWaV278 durch Bistolas et al. (2017) deutete auf eine Zugehörigkeit zur ersten Gruppe, insbesondere ist das CAP homolog zu dem von CRUV-15-B. Es ist mit 3478 nt (Nukleotiden oder Basen) wie zu erwarten größer als das der meisten CRESS-DNA-Viren, aber in dieser Größe vergleichbar mit dem der anderen Mitgliedern der Cruciviren-Kladen.[8]
IWaV278 zeigte keine Anzeichen von Intra-Gen-Chimärismus (en. intra-gene chimerism – mehrfachen Rekombinationsereignissen innerhalb des REP-Gens) oder teilweisem Austausch von REP-Genen. Derartige Phänomene hat man allerdings bei mehreren anderen Cruciviridae-Genomen gefunden.[8]
Weitere Forschungsergebnisse
BearbeitenEine neuere noch umfangreichere Analyse von Higuera et al. (2020) zeigte aber, dass die REPs der Cruciviren die Vielfalt aller anderen CRESS-DNA-Viren überspannt und die Grenzen zwischen ihnen verwischt. Es gibt Cruciviren, die bzgl. des REP eng mit Geminivirdae und Genomoviridae (den beiden Familien der Ordnung Geplafuvirales) verwandt sind, also (einigermaßen) in das obige Schema passen. Man fand aber auch Cruciviren mit REP-Homologie zu den anderen klassifizierten (d. h. offiziellen, vom International Committee on Taxonomy of Viruses (ICTV) bestätigten) CRESS-DNA-Familien. Interessanterweise enthält der Sonderfall CruV-420 nicht ein, sondern zwei verschiedene tombusvirenartige Kapsidproteine.[5] Insgesamt deuteten diese Ergebnisse darauf hin, dass Rekombination tendenziell häufiger zwischen Gruppen von Cruciviren mit relativ ähnlichen Kapsidproteinen auftrat (oder auftritt).[5]
Beschreibung
BearbeitenAufbau
BearbeitenSolange alle vorgeschlagenen Vertreter der Cruciviren aus Metagenomanalysen stammen, lassen sich noch keine sicheren Aussagen über die Morphologie machen. Es wurden zwar EM-Aufnahmen virusartiger Partikel (en. virus-like particles) gemacht.[37] Es bleibt daher nachzuweisen, dass diese Träger der Genome aus der Metagenomik, d. h. die gesuchten Virionen (Virusteilchen) sind.
Genom
Bearbeiten.png)
Die Balken unter den ORFs zeigen von der Analyse erkannte Proteinfamilien an.[4]
.jpg)
grün: vorhergesagte Kapsidprotein-Gene (CAP);
rot: vorhergesagte Replikations-Initiator-Protein-Gene (REP);[Anm. 13]
lila: mutmaßlicher Replikationsursprung.
Hintergrund: Das Entnahmegebiets der Proben.
Unten rechts Karte mit Lage von Woodburn (Stern).[20]
Genomkarten von CHIV1 bis CHIV13 finden sich bei Roux et al. (2013) Fig. 1 (inklusive Längen).[1]
Proteom
BearbeitenEinige Beispiele für vorhergesagte Proteinstrukturen:
![Vorhergesagte REP-Struktur von BSL-RDHV[4]](/wiki/Datei:13062_2012_Article_325_Fig7_HTML-BSL-RDHV-CP%2BREP-Structure-(B).jpg)
![Vorhergesagte CAP-Struktur von BSL-RDHV im Vergleich zu der von TBSV und MNSV[4]](/wiki/Datei:13062_2012_Article_325_Fig7_HTML-BSL-RDHV-CP%2BREP-Structure-(A).png)
![CAP-Strukturmodell von CruV-359[5]](/wiki/Datei:F2.large-cruciviruses-ProteinConservation-A(btm).jpg)
![CAP-Strukturmodell von CHIV14[18]](/wiki/Datei:2021_03_25_18_03_51_evv034_Fig2C-CHIV14-CP.png)
Replikation
BearbeitenFast alle ssDNA-Viren mit zirkulärem Genom replizieren dieses auf ähnliche Weise (sog. Rolling Circle Replication, RCR). Zunächst wird zur ssDNA das zugehörige Komplement synthetisiert (der Wirt hat die nötigen Enzyme für seine eigene Replikation), so dass eine (zirkuläre) Doppelstrang-DNA (dsDNA) entsteht.[38] Das REP-Protein startet (initiiert) dann die Replikation, indem es die zirkuläre dsDNA an der konservierten Haarnadelstruktur (en. stem-loop structure), dem Doppelstrangursprung (DSO), aufschneidet. Dies liefert einen Startpunkt (en. Primer) für die DNA-Replikation vermöge der DNA-Polymerasen der Wirtszelle. Das Rep-Protein in den meisten ssDNA-Viren ist in einer Nebenfunktion auch eine Helikase und schält den Komplementärstrang der DNA wieder ab. Schließlich fügt das REP-Protein das Genom nach einer einzigen Replikationsrunde wieder zusammen (Ligation).[2]
Bedeutung
BearbeitenCruciviren sind von entscheidender Bedeutung für das Verständnis der Virusevolution und der Entstehung neuer Viren durch Rekombination und Hybridisierung. Sie können neue Einblicke in den Übergang von RNA zu DNA in der Evolution des frühen zellulären Lebens auf der Erde geben (RNA-Welt-Hypothese).[2][1]
Die das REP kodierenden Gene (genauer: Offener Leserahmen, englisch open reading frames, ORFs) der Cruciviren haben die größte Homologie zu Einzelstrang-DNA-Viren (ssDNA-Viren, Baltimore-Gruppe 2), während die das Kapsidprotein kodierenden Gene (ORFs) die meiste Homologie zu Einzelstrang-RNA-Viren (ssRNA-Viren, Baltimore-Gruppe 4 und 5) haben.
Während das Genom zellulärer Organismen stets aus einem oder mehreren Molekülen Doppelstrang-DNA (dsDNA) besteht, sind Viren die einzigen rezenten Organismen (bzw. molekulare Maschinen), die neben DNA auch RNA als genetisches Material verwenden, sowohl als Einzel- als auch als Doppelstrang.[2]
Zwar tauschen alle Viren innerhalb von Gruppen mehr oder weniger untereinander verwandter Viren genetisches Material aus, d. h. insbesondere DNA-Viren mit DNA-Viren und RNA-Viren mit RNA-Viren. Ein Austausch genetisches Material zwischen RNA- und DNA-Viren war aber bis zur Entdeckung der RNA-DNA-Hybridviren (RDHVs) nicht beobachtet worden.[2] Dies ist um so erstaunlicher, als die (nach den Annehmen) den Cruciviren zugrunde liegende Hybridisierung eine umgekehrte (reverse) Transkription voraussetzt (siehe Reverse Transkriptase), die normale Transkriptionsrichtung ist DNA → RNA (mRNA oder tRNA).
Im Rahmen der RNA-Welt-Hypothese wird vermutet, dass Viren die ersten waren, die Doppelstrang-DNA (dsDNA) als genetisches Material in einer vermuteten primordialen (ursprünglichen) RNA-Welt verwendet haben, und dass diese Strategie dann vom zellulären Leben (zunächst organisiert in hypothetischen Ribozyten alias Ribozellen) übernommen wurde. Daher könnten RNA-Viren das Überbleibsel dieser RNA-Welt sein; RHDVs könnten dagegen moderne Relikte aus der Übergangszeit sein – ähnlich wie es von Retroviren (wie die HIVs, Baltimore-Gruppe 6) und Pararetroviren (wie die Cacao-swollen-shoot-Viren, CSSVs, Baltimore-Gruppe 7) vermutet wird – diese wechseln als Teil ihres Replikationszyklus zwischen DNA und RNA.[2] Auch bei den Retroviren wurde bereits 1970 eine Reverse Transkriptase gefunden.[39][40]
Das offensichtlich von einem RNA-Virus erworbene Gen für das Kapsidprotein bestimmt die äußere Hülle der Viruspartikel (Virionen) – und damit aber auch, welche Wirte das Virus infizieren kann, wie es übertragen wird und wie es die Wirtsabwehr umgeht. Offenbar hat dieser Erwerb den Cruciviren auf so einen evolutionären Vorteil verschafft, weshalb sich die Hybride so weit kosmopolitisch etablieren konnten.[2]
Anmerkungen
Bearbeiten- ↑ aus 9 Basen bestehend
- ↑ Die Begriffen „hybrid“ oder „chimär“ dürfen nicht so verstanden werden, als ob das Genom chimär teils aus RNA und teils aus RNA bestünde, beide Begriffe beziehen sich lediglich auf die RNA-Herkunft eines Teils des DNA-Genoms.
- ↑ Bei der beim National Center for Biotechnology Information (NCBI) als Gattung geführte Name „Crucivirus“ handelt es sich möglicherweise lediglich um die Einzahlform der informellen (nicht-taxonomischen) Bezeichnung Cruciviruses. Eine nähere Verwandtschaft als bei den anderen unter „Cruciviridae“ geführten Kandidaten offenbar nirgend s beschrieben. Insbesondere fällt auf, dass das NCBI BSL-RDHV zwar listet, aber weder unter einer „Gattung“ „Crucivirus“ noch unter der vorgeschlagenen Familie „Cruciviridae“; für den zuerst gefundenen Vertreter wäre dies als Typusart aber zu erwarten. Möglicherweise ist dieser Umstand historisch bedingt.
- ↑ NCBI: nicht in Familie „Cruciviridae“
- ↑ a b NCBI: in „Gattung“ „Crucivirus“
- ↑ verschrieben als CHIV4 bei Bistolas et al., 2017, die Zugriffsnummer YP_009142777.1 ist korrekt
- ↑ englisch peat bog ‚Torfmoor‘; marsh ‚Sumpf‘.
- ↑ a b Möglicherweise ist PB1-RDHV identisch mit CruV-MC1, man beachte die Genomlängen.
- ↑ F=Fenn: Nieder- oder Flachmoor (en. Fen, B=(Hoch-)Moor (en. Bog)
- ↑ Bei Virussequenzen aus der Metagenomik ist oft zunächst unbekannt, welche Wirte diese Viren infizieren. Infrage kommen als Wirte solche Organismen, deren Genom zusammen mit den betrachteten Virussequenzen gefunden wird. Offenbar sind diese Organismen und die Viren dann in irgendeiner Form vergesellschaftet („assoziiert“). Das muss nicht notwendig bedeuten, dass die Viren diese Organismen als Wirte parasitieren, evtl. werden diese lediglich als Vektoren benutzt, oder die Viren infizieren ihrerseits Parasiten dieser Organismen.
- ↑ Die sog. Eipilze sind keine Pilze, sondern Verwandte der Braun- und Goldalgen.
- ↑ Aus demselben Grund könnte die bisherige Taxonomie der CRESS-DNA-Viren in die bisherigen Familien infrage gestellt werden, nämlich in dem Maße, als diese auf Basis von REP-Homologien erfolgt ist.
- ↑ siehe Porcines Circovirus-1 (PCV1)
Weblinks
Bearbeiten- Mart Krupovic: Recombination between RNA viruses and plasmids might have played a central role in the origin and evolution of small DNA viruses, in: BioEssays, 7. August 2012, doi:10.1002/bies.201200083
- Mart Krupovic, Arvind Varsani, Jens H. Kuhn, Darius Kazlauskas, Mya Breitbart, E. Delwart, K. Rosario, N. Yutin, Y. I. Wolf, B. Harrach, F. M. Zerbini, V. V. Dolja, E. V. Koonin: [1] (docx, xlsx), Proposal to ICTV (accepted): Cressdnaviricota
- Mart Krupovic, Arvind Varsani, Darius Kazlauskas, Mya Breitbart, Eric Delwart, Karyna Rosario, Natalya Yutin, Yuri I. Wolf, Balázs Harrach, F. Murilo Zerbini, Valerian V. Dolja, Jens H. Kuhn, Eugene V. Koonin; Rozanne M. Sandri-Goldin (Hrsg.): Cressdnaviricota: a Virus Phylum Unifying Seven Families of Rep-Encoding Viruses with Single-Stranded, Circular DNA Genomes, in: ASM Journal of Virology Band 94, Nr. 12, 1. Juni 2020, e00582-20; doi:10.1128/JVI.00582-20: Cressdnaviricota
- Vincent Racaniello: A DNA virus with the capsid of an RNA virus, auf: virology blog vom 19. Juli 2012: BSL-RDHV
- Karyna Rosario, Milen Marinov, Daisy Stainton, Simona Kraberger, Elizabeth J. Wiltshire, David A. Collings, Matthew Walters, Darren P. Martin, Mya Breitbart, Arvind Varsani: Dragonfly cyclovirus, a novel single-stranded DNA virus discovered in dragonflies (Odonata: Anisoptera), in: J Gen Virol 92(pt 6), Juni 2011, S. 1302–1308, Epub 2. März 2011, doi:10.1099/vir.0.030338-0, PMID 21367985.
- Kenneth M. Stedman: Deep Recombination: RNA and ssDNA Virus Genes in DNA Virus and Host Genomes, in: Annual Review of Virology, Band 2, 2. September 2015, S. 203–217, doi:10.1146/annurev-virology-100114-055127
Einzelnachweise
Bearbeiten- ↑ a b c d e f g h i j k l m n o p q r Simon Roux, François Enault, Gisèle Bronner, Daniel Vaulot, Patrick Forterre, Mart Krupovic: Chimeric viruses blur the borders between the major groups of eukaryotic single-stranded DNA viruses, in: Nat Commun 4:2700, 6. November 2013, doi:10.1038/ncomms3700. Siehe insbesondere Supplement 2.
- ↑ a b c d e f g h i j k l m n o p Ignacio (Nacho) de la Higuera, Ellis Torrance, Amber Maluenda, George Kasun, Max Larson; Rita Clare: Cruciviruses / RNA-DNA Hybrid Viruses, Extreme Virus Lab (Stedman Lab), 2018. Insbesondere Fig. Cruciviridae
- ↑ a b c Richard Harth: Criss-crossing viruses give rise to peculiar hybrid variants ( des vom 5. August 2021 im Internet Archive) Info: Der Archivlink wurde automatisch eingesetzt und noch nicht geprüft. Bitte prüfe Original- und Archivlink gemäß Anleitung und entferne dann diesen Hinweis., Arizona State University, Biodesign Institute, 29. Oktober 2020; sowie
Cruciviruses: Criss-Crossing Viruses Give Rise to Peculiar Hybrid Variants, auf: SciTechDaily vom 2. November 2020. Siehe insbes. Cruciviruses ( des vom 2. November 2021 im Internet Archive) Info: Der Archivlink wurde automatisch eingesetzt und noch nicht geprüft. Bitte prüfe Original- und Archivlink gemäß Anleitung und entferne dann diesen Hinweis. (Harth, Biodesign), alternativ: Cruciviruses (SciTechDaily) - ↑ a b c d e Geoffrey S. Diemer, Kenneth M. Stedman: A novel virus genome discovered in an extreme environment suggests recombination between unrelated groups of RNA and DNA viruses, auf: PDXScholar, University Library, Portland State University, 6. Dezember 2012. Ebenso in: Biol Direct. Band 7, Nr. 13, 11. Juni 2012, doi:10.1186/1745-6150-7-13, PMID 22515485, PMC 3372434 (freier Volltext)
- ↑ a b c d e f g Ignacio (Nacho) de la Higuera, George W. Kasun, Ellis L. Torrance, Alyssa A. Pratt, Amberlee Maluenda, Jonathan Colombet, Arvind Varsani, Kenneth M. Stedman et al.; Stephen J. Giovannoni (Hrsg.): Unveiling Crucivirus Diversity by Mining Metagenomic Data, in: ASM mBio Band 11, Nr. 5, e01410-20; 1. September 2020; doi:10.1128/mBio.01410-20, PMID 32873755, PMC 7468197 (freier Volltext).
- ↑ Olivia Steel, Simona Kraberger, Alyssa Sikorski, Laura M.Young, Ryan J. Catchpole, Aaron J. Stevens, Jenny J. Ladley, Dorien S. Coraya, Daisy Stainton, Anisha Dayaram, Laurel Julian, Katherine van Bysterveldt, Arvind Varsani: Circular replication-associated protein encoding DNA viruses identified in the faecal matter of various animals in New Zealand, in: Genetics and Evolution 43, September 2016, S. 151–164, doi:10.1016/j.meegid.2016.05.008
- ↑ Anisha Dayarama, Mark L. Galatowitsch, Gerardo R. Argüello-Astorg, Katherine van Bysterveldt, Simona Kraberger, Daisy Stainton, Jon S. Harding, Philippe Roumagnac, Darren P. Martin, Pierre Lefeuvre, Arvind Varsani: Diverse circular replication-associated protein encoding viruses circulating in invertebrates within a lake ecosystem, in: Infection, Genetics and Evolution 39, April 2016, Epub 10. Februar 2016, S. 304–316. doi:10.1016/j.meegid.2016.02.011
- ↑ a b c d e f g h Kalia S. I. Bistolas, Ryan M. Besemer, Lars G. Rudstam, Ian Hewson: Distribution and Inferred Evolutionary Characteristics of a Chimeric ssDNA Virus Associated with Intertidal Marine Isopods, in: MDPI Viruses Band 9, Nr 12, 361; Special Issue Viral Recombination: Ecology, Evolution and Pathogenesis; Dezember 2017, Epub 26. November 2017; doi:10.3390/v9120361, PMID 29186875, PMC 5744136 (freier Volltext).
- ↑ NCBI: Boiling Springs Lake RNA-DNA hybrid virus (species)
- ↑ NCBI: Idotea virus IWaV278 (species)
- ↑ Pfam: Proteome: Idotea virus IWaV278 (NCBI tax. ID 2058759)
- ↑ UniProt: Idotea virus IWaV278 (SPECIES)
- ↑ NCBI: RDHV-like virus SF1 (species), AKM12421.1
- ↑ Karyna Rosario, Kaitlin Mettel, Bayleigh Benner, Ryan Johnson, Catherine Scott, Sohath Yusseff-Vanegas, Christopher Baker, Deby Cassill, Caroline Storer, Arvind Varsani, Mya Breitbart: Virus Discovery in All Three Major Lineages of Terrestrial Arthropods Highlights the Diversity of Single-stranded DNA Viruses Associated with Invertebrates, University of South Florida, College of Marine Science: Scholar Commons — Marine Science Faculty Publications, 2017, 652. Part of the Life Sciences Commons. 11. Oktober 2018 PeerJ, PMID 30324030, PMC 6186406 (freier Volltext), doi:10.7717/peerj.5761
- ↑ NCBI: Fire ant associated circular virus 1 (species)
- ↑ NCBI: Sewage-associated circular DNA virus-13 (species, 664651386)
- ↑ a b c Achim Quaiser, Mart Krupovic, Alexis Dufresne, André-Jean Francez, Simon Roux: Diversity and comparative genomics of chimeric viruses in Sphagnum-dominated peatlands, in: Virus Evolution, Band 2, Nr. 2, Juli/Oktober 2016, vew025, doi:10.1093/ve/vew025
- ↑ a b Mart Krupovic, Ning Zhi, Jungang Li, Gangqing Hu, Eugene V. Koonin, Susan Wong, Sofiya Shevchenko, Keji Zhao, Neal S. Young: Multiple Layers of Chimerism in a Single-Stranded DNA Virus Discovered by Deep Sequencing, in: Genome Biol. Evol. (GBE) Band 7, Nr. 4, 16. Februar 2015, S. 993–1001, doi:10.1093/gbe/evv034
- ↑ NCBI: Chimeric virus 14 (species)
- ↑ a b Ignacio (Nacho) de la Higuera, Ellis Lynn Torrance, Alyssa A. Pratt, George W. Kasun, Amberlee Maluenda, Kenneth M. Stedman: Genome Sequences of Three Cruciviruses Found in the Willamette Valley (Oregon), in: Microbiology Resource Announcements Band 8, Nr. 23, 6. Juni 2019, e00447-19; doi:10.1128/MRA.00447-19, PMID 31171623, Europe PMC MED/31171623, PMC 6554611 (freier Volltext)
- ↑ NCBI: unclassified Crucivirus (list) bzw. Crucivirus*-##* (list)
- ↑ NCBI: Crucivirus-like circular genetic element-85 (species)
- ↑ NCBI: Crucivirus-like circular genetic element-471 (species)
- ↑ NCBI: Dragonfly cyclicusvirus (species, unclassified ssDNA)
- ↑ a b Karyna Rosario, Anisha Dayaram, Milen Marinov, Jessica Ware, Simona Kraberger, Daisy Stainton, Mya Breitbart, Arvind Varsani: Diverse circular ssDNA viruses discovered in dragonflies (Odonata: Epiprocta), in: Journal of General Virology Band 93, Nr. 12, 1. Dezember 2012, doi:10.1099/vir.0.045948-0. Insbes. Tbl. 2
- ↑ NCBI: Satellite tobacco mosaic virus (no rank)
- ↑ NCBI: Virtovirus (species)
- ↑ Eugene V. Koonin, Valerian V. Dolja: Expanding networks of RNA virus evolution, In: BMC Biology Band 10, Nr. 54, 20. Juni 2012, doi:10.1186/1741-7007-10-54
- ↑ Xiao Yu, Bo Li, Yanping Fu, Daohong Jiang, Said A Ghabrial, Guoqing Li, Youliang Peng, Jiatao Xie, Jiasen Cheng, Junbin Huang, Xianhong Yi: A geminivirus-related DNA mycovirus that confers hypovirulence to a plant pathogenic fungus, in: PNAS USA 107(18). 4. Mai 2010, S. 8387–8392, doi:10.1073/pnas.0913535107, PMID 20404139
- ↑ Darius Kazlauskas, Anisha Dayaram, Simona Kraberger, Sharyn Goldstien, Arvind Varsani, Mart Krupovic: Evolutionary history of ssDNA bacilladnaviruses features horizontal acquisition of the capsid gene from ssRNA nodaviruses. In: Virology. 504. Jahrgang, 2017, S. 114–121, doi:10.1016/j.virol.2017.02.001, PMID 28189969 (sciencedirect.com).
- ↑ Mart Krupovic, Eugene V. Koonin: Evolution of eukaryotic single-stranded DNA viruses of the Bidnaviridae family from genes of four other groups of widely different viruses, in: Nature Sci Rep Band 4, Nr. 5347, 18. Juni 2014, doi:10.1038/srep05347. Siehe insbes. Fig. 1
- ↑ a b Michael J. Tisza, Diana V. Pastrana, Nicole L. Welch, Brittany Stewart, Alberto Peretti, Gabriel J. Starrett, Yuk-Ying S. Pang, Siddharth R. Krishnamurthy, Patricia A. Pesavento, David H. McDermott, Philip M. Murphy, Jessica L. Whited, Bess Miller, Jason Brenchley, Stephan P. Rosshart, Barbara Rehermann, John Doorbar, Blake A. Ta'ala, Olga Pletnikova, Juan C. Troncoso, Susan M. Resnick, Ben Bolduc, Matthew B. Sullivan, Arvind Varsani, Anca M. Segall, Christopher B. Buck: Discovery of several thousand highly diverse circular DNA viruses, in: eLife 9, 4. Februar 2020, e51971; doi:10.7554/eLife.51971. Siehe insbes. Fig. 4
- ↑ NCBI: Sclerophthora macrospora virus A (species), (+)ssRNA-Virus
- ↑ NCBI: Plasmopara halstedii virus A (species), Plasmopara halstedii virus A isolate Ph4-93 RNA-dependent RNA polymerase (RdRp) gene, partial cds, RNA-Virus
- ↑ a b ICTV: ICTV Master Species List 2019.v1, New MSL including all taxa updates since the 2018b release, March 2020 (MSL #35)
- ↑ NCBI: Cruciviridae (family)
- ↑ Ian Hewson, Gabriel Ng, WenFang Li, Brenna A. LaBarre, Isabel Aguirre, Jorge G. Barbosa, Mya Breitbart, Anthony W. Greco, Colleen M. Kearns, Alexander Looi et al.: Metagenomic identification, seasonal dynamics, and potential transmission mechanisms of a Daphnia‐associated single‐stranded DNA virus in two temperate lakes, in: ASLO Limnology And Oceanography Band 58, Nr. 5, September 2013, Epub 28. Juli 2013, S. 1605–1620, doi:10.4319/lo.2013.58.5.1605
- ↑ SIB: ssDNA Rolling circle. In: ViralZone. Swiss Institute of Bioinformatics (SIB): Expasy, abgerufen am 29. März 2021.
- ↑ H. M. Temin, S. Mizutani: RNA-dependent DNA polymerase in virions of Rous sarcoma virus. In: Nature. Band 226, Nr. 5252, Juni 1970, S. 1211–1213, PMID 4316301.
- ↑ D. Baltimore: RNA-dependent DNA polymerase in virions of RNA tumour viruses. In: Nature. Band 226, Nr. 5252, Juni 1970, S. 1209–1211, PMID 4316300.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}